Love may be in the air on Valentine’s day, but sweat, urine, and other secreted bodily fluids fill the other 364 days of the year. Best known to increase attraction and alter moods, ‘ectohormone’ refers to the intraspecific chemicals that affect the activity of other individuals through the expulsion of biofluids (Doty, 2014). Since discovery, ectohormone evolved into a series of synonymous terms before settling in 1959 to ‘pheromone’ a widely recognizable substance capable of triggering a social response. Differing from the internal communication of hormones, pheromones work outside the body to target members within the same species. It aids in attraction, hunting, and recognition through the vomeronasal organ, a tubular structure located in the anterior of the nasal cavity. Although this susceptibility of scents is prevalent to humans, insects were first discovered exhibiting similar adaptations (Verhaeghe, Gheysen and Enzlin, 2013).
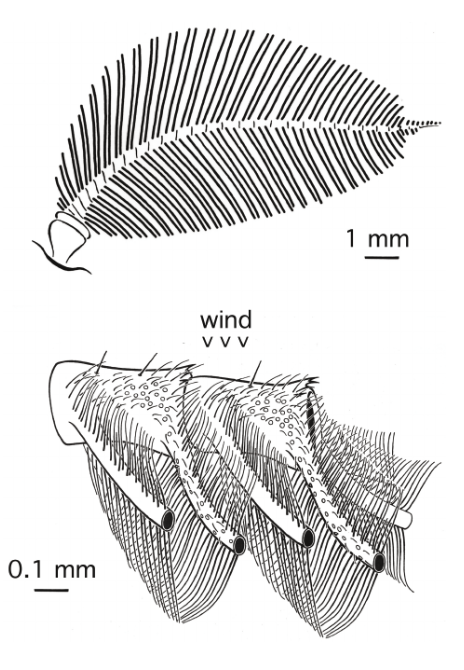
Many male moths have enlarged antennae containing sensilla, sensory hairs or plates involved in nerve connections as seen in Figure 1, serving the purpose of detection and recognition of specific stimuli. Female Saturniid moths utilize the sensilla of their male counterparts, and are reported to detect the pheromones over distances of one kilometer using an antenna with an area comparable to the size of a human nostril (Kaissling, 2014).

Figure 1: Enlarged diagram of a male Saturniid moth with four distinct sides of the antennal stem segment. Each olfactory hair contains two or three receptor neurons responding to two or three components of the female pheromone (Kaissling, 2009).
Other moths commonly known to employ the olfactory effects of pheromones are female silkworms. Secreting a volatile sixteen carbon alcohol, bombykol attracts male silkworms through the antenna, as it contains a pheromone-binding protein that conveys pheromones to a membrane-bound receptor on the nerve cell (Sandler et al., 2000). Bombykol is now recognized as the first pheromone to be chemically characterized, but is far from being the only one attributing to sexual desire.
Surpassing the boundaries of natural life, robotic insects have captured the interest of the swarm intelligence field as being a system of programmable 4 centimeter automations, mimicking ant and bee colony optimization. They are used in micromachinery within the human body, mining exploration, and even disaster rescue missions. Swarm robots are capable of deploying both attractive and repellent pheromone based chemicals from the artificial platform ColCOS Φ in furthering scientific research on pheromone stimulation, and social structure of insects (Liu et al., 2020). Modelled by the following equation, Figure 2 represents the source position and strength of the pheromone to determine the diffusion effect of chemicals given no wind or external disturbances.

Figure 2: Displayed in a differential equation with evaporation and diffusion parameters on the assumption that the source is continuously secreting the pheromone. (μx,μy) defines the source of the pheromone, K defines the strength of the pheromone, and σx and σy are the standard deviation of the Gaussian function which determines the effectiveness of diffusion (Liu et al., 2020).
Upon initial release, swarm robots will first enter ‘wandering mode’ as they begin to explore the surrounding area in a random-like manner. Agents will continuously explore the environment until confronted with a boundary or obstacle, before an ‘avoiding collision’ mode will override the controller, and avoid the looming encounter. This heedless wander will continue, until met with a diffused cloud of pheromones secreted by other swarm robots. Once entered, it will trigger ‘pheromone guide motion’, a mode that senses if the value of the attractive pheromone is greater than the programmed threshold, where the agent can stop and start to begin complex coordinated functions (Liu et al., 2020).
Using pheromones, humans, moths, and robots, have the potential for increased communication, delving into the hierarchy of social interactions. Accelerating all we think we know about the intraspecifics of chemical signalling, and all we have yet to discover.
References
Doty, R.L., 2014. Human Pheromones: Do They Exist? In: C. Mucignat-Caretta, ed. Neurobiology of Chemical Communication, Frontiers in Neuroscience. Boca Raton (FL): CRC Press/Taylor & Francis. Available at: http://www.ncbi.nlm.nih.gov/books/NBK200980/ [Accessed 22 January 2024].
Kaissling, K.-E., 2009. The Sensitivity of the Insect Nose: The Example of Bombyx Mori. In: Studies in Computational Intelligence. pp.45–52. https://doi.org/10.1007/978-3-642-00176-5_3.
Kaissling, K.-E., 2014. Pheromone Reception in Insects: The Example of Silk Moths. In: C. Mucignat-Caretta, ed. Neurobiology of Chemical Communication, Frontiers in Neuroscience. Boca Raton (FL): CRC Press/Taylor & Francis. Available at: http://www.ncbi.nlm.nih.gov/books/NBK200991/ [Accessed 23 January 2024].
Liu, T., Sun, X., Hu, C., Fu, Q., Isakhani, H. and Yue, S., 2020. Investigating Multiple Pheromones in Swarm Robots – A Case Study of Multi-Robot Deployment. In: 2020 5th International Conference on Advanced Robotics and Mechatronics (ICARM). 2020 5th International Conference on Advanced Robotics and Mechatronics (ICARM). pp.595–601. https://doi.org/10.1109/ICARM49381.2020.9195311.
Sandler, B.H., Nikonova, L., Leal, W.S. and Clardy, J., 2000. Sexual attraction in the silkworm moth: structure of the pheromone-binding-protein-bombykol complex. Chemistry & Biology, 7(2), pp.143–151. https://doi.org/10.1016/s1074-5521(00)00078-8.
Verhaeghe, J., Gheysen, R. and Enzlin, P., 2013. Pheromones and their effect on women’s mood and sexuality. Facts, Views & Vision in ObGyn, 5(3), pp.189–195. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3987372/ [Accessed 25 January 2024].