Mitochondrial DNA (mtDNA) is inherited solely from the mother, making it useful in tracing maternal lineage and studying human evolution. Its resilience in damaged samples helps track population movements across continents, revealing our common ancestor, “Mitochondrial Eve.”
It is theorized that the mitochondria were once a prokaryotic microbe that was engulfed by a eukaryote (Martin, Garg and Zimorski, 2015). The development of this endosymbiotic relationship allowed for facilitated habitation in aerobic conditions. However, their similarities to prokaryotic DNA includes the lack of protective histones, limited proofreading and repair mechanisms (with an increased risk of mutations 10 to 20 times compared to the nuclear genome), as well as the generation of reactive oxidizing species from mitochondrial function (Lax, Turnbull and Reeve, 2011).
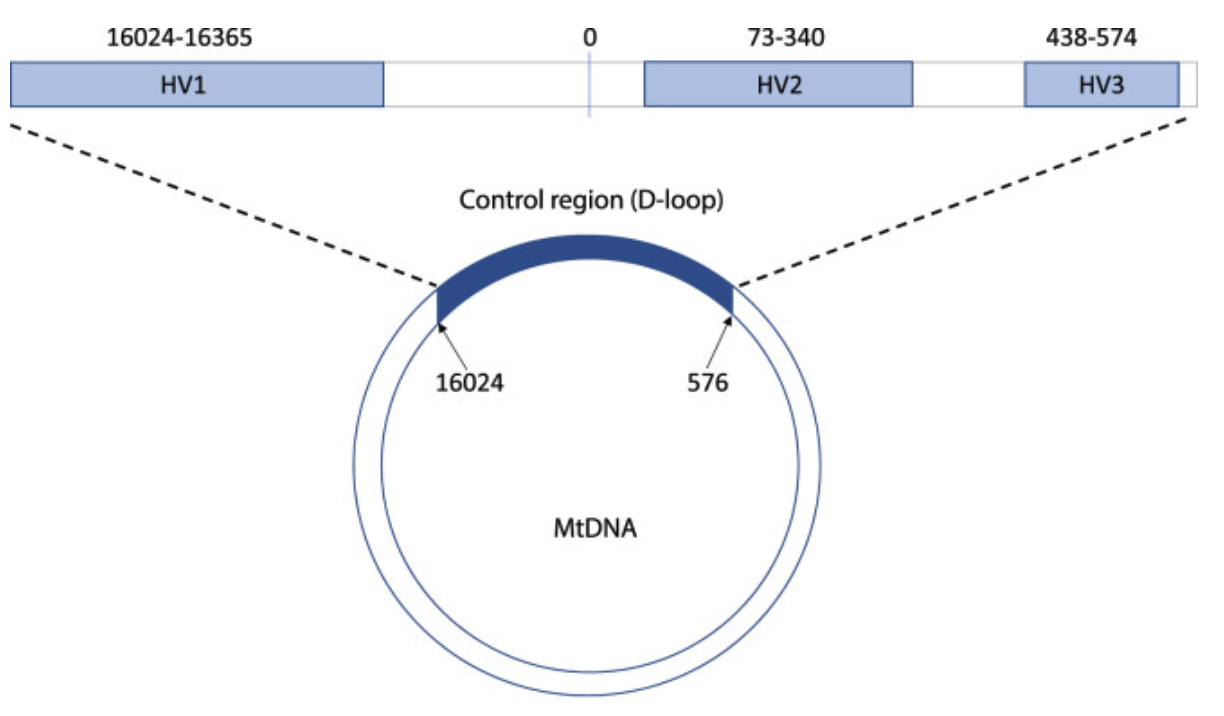
The standard mtDNA genome for comparison today would be the revised Cambridge Reference Sequence (rCRS) where each nucleotide is numbered (Bandelt, et al., 2014). The displacement (D) loop or control region which is 1,124 bp in size, spans from 16,024-576 in position, is a non-coding region that contains needed transcription and replication elements (Figure 1) (Sharma, et al., 2005). The highest volume of sequence variability in the genome is found in three sites of the control region, which are known as the hypervariable (HV) regions (Syndercombe Court, 2021). HV1 covers nucleotides 16,024 to 16,365, HV2 from 73 to 340 and HV3 from 438 to 574. Greater variability is found in HV1 and HV2 as compared to HV3, making them frequently sequenced in genetic analysis.

The high rate of mutations in mtDNA would result in heteroplasmy (variation of genotypes) in the population that is transmitted through the genetic bottleneck effect (Parakatselaki and Ladoukakis, 2021). The wild-to-variant balance can change over time due to genetic drift and agnostic (beneficial) expression can become homoplasmic (fixed) in the local population. The major haplogroups or varying mitochondrial types in distinct global populations were labelled from A to Z in association with potential migration routes of Homo sapiens (Figure 2) (Syndercombe Court, 2021). The most recent common ancestor to humans today is referred to as “Mitochondrial Eve,” who was present about 120,000 to 156,000 years ago. She was the predecessor who had unbroken matrilineal succession through her female offspring, without being the first or only woman of the species.

Migration from present-day Serbia in groups M as well as A,C, and D established the indigenous populations of North America about 16,000 years ago (Wooller, et al., 2018). This was due to the theorized crossing of the Bering Land Bridge that was present under lower sea levels. The migration from North America to South America was a result of crossing the Isthmus of Panama which was formed about 2.8 million years ago (O’Dea, et al., 2016).
Applications of mtDNA sequencing allow for the detection of variability, which can be used to lineate the evolution of humans through maternal succession in haplogroups. Through further improvement of mtDNA sequencing, its selective use can be expanded to improve forensic science as well as medical therapy practices, and further anthropological analysis through genetic understanding.
References:
Bandelt, H.-J., Kloss-Brandstätter, A., Richards, M.B., Yao, Y.-G. and Logan, I., 2014. The case for the continuing use of the revised Cambridge Reference Sequence (rCRS) and the standardization of notation in human mitochondrial DNA studies. Journal of Human Genetics, 59(2), pp.66–77. https://doi.org/10.1038/jhg.2013.120.
Lax, N.Z., Turnbull, D.M. and Reeve, A.K., 2011. Mitochondrial Mutations. The Neuroscientist, 17(6), pp.645–658. https://doi.org/10.1177/1073858411385469.
Martin, W.F., Garg, S. and Zimorski, V., 2015. Endosymbiotic theories for eukaryote origin. Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1678), p.20140330. https://doi.org/10.1098/rstb.2014.0330.
O’Dea, A., Lessios, H.A., Coates, A.G., Eytan, R.I., Restrepo-Moreno, S.A., Cione, A.L., Collins, L.S., de Queiroz, A., Farris, D.W., Norris, R.D., Stallard, R.F., Woodburne, M.O., Aguilera, O., Aubry, M.-P., Berggren, W.A., Budd, A.F., Cozzuol, M.A., Coppard, S.E., Duque-Caro, H., Finnegan, S., Gasparini, G.M., Grossman, E.L., Johnson, K.G., Keigwin, L.D., Knowlton, N., Leigh, E.G., Leonard-Pingel, J.S., Marko, P.B., Pyenson, N.D., Rachello-Dolmen, P.G., Soibelzon, E., Soibelzon, L., Todd, J.A., Vermeij, G.J. and Jackson, J.B.C., 2016. Formation of the Isthmus of Panama. Science Advances, 2(8), p.e1600883. https://doi.org/10.1126/sciadv.1600883.
Parakatselaki, M.-E. and Ladoukakis, E.D., 2021. mtDNA Heteroplasmy: Origin, Detection, Significance, and Evolutionary Consequences. Life, 11(7), p.633. https://doi.org/10.3390/life11070633.
Sharma, H., Singh, A., Sharma, C., Jain, S.K. and Singh, N., 2005. Mutations in the mitochondrial DNA D-loop region are frequent in cervical cancer. Cancer Cell International, 5, p.34. https://doi.org/10.1186/1475-2867-5-34.
Syndercombe Court, D., 2021. Mitochondrial DNA in forensic use. Emerging Topics in Life Sciences, 5(3), pp.415–426. https://doi.org/10.1042/ETLS20210204.
Wooller, M.J., Saulnier-Talbot, É., Potter, B.A., Belmecheri, S., Bigelow, N., Choy, K., Cwynar, L.C., Davies, K., Graham, R.W., Kurek, J., Langdon, P., Medeiros, A., Rawcliffe, R., Wang, Y. and Williams, J.W., 2018. A new terrestrial palaeoenvironmental record from the Bering Land Bridge and context for human dispersal. Royal Society Open Science, 5(6), p.180145. https://doi.org/10.1098/rsos.180145.